Мутагенные факторы и мутагенез
Содержание:
Глава 4. ОРГАНИЗМЕННЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ЖИВОГО
4.11. МУТАГЕННЫЕ ФАКТОРЫ
Любые мутации могут возникнуть спонтанно или быть индуцированными. Спонтанные мутации появляются под влиянием неизвестных природных факторов и приводят к ошибкам при репликации ДНК.
Индуцированные мутации возникают под воздействием специальных направленных факторов, повышающих мутационный процесс.
Мутагенным действием обладают факторы физической, химической и биологической природы.
Среди физических мутагенов наиболее сильное мутантное действие оказывает ионизирующая радиация — рентгеновские лучи, α-, β-, γ-лучи. Обладая большой проникающей способностью, при действии на организм они вызывают образование свободных радикалов ОН или НО2 из воды, находящейся в тканях. Эти радикалы обладают высокой реакционной способностью. Они могут расщеплять нуклеиновые кислоты и другие органические вещества.
Облучение вызывает как генные, так и хромосомные перестройки.
Ультрафиолетовое излучение характеризуется меньшей энергией, не вызывающей ионизацию тканей. Действие УФ-излучения приводит к образованию тимидиновых димеров. Присутствие димеров в ДНК приводит к ошибкам при ее репликации.
Химические мутагены должны обладать следующими качествами:
• высокой проникающей способностью;
• свойством изменять коллоидное состояние хромосом;
• определенным действием на состояние хромосомы или гена. К химическим веществам, вызывающим мутации, можно отнести органические и неорганические вещества, такие, как кислоты, щелочи, перекиси, соли металлов, формальдегид, пестициды, дефолианты, гербициды, колхицин и др.
Некоторые вещества способны усиливать мутационный эффект в сотни раз по сравнению со спонтанным. Их называют супермутагенами. Эти супермутагены вызывают широкий спектр точковых мутаций в концентрациях меньше тех, которые индуцируют хромосомные перестройки, видимые под микроскопом. Супермутагенной активностью обладают нитрозосоединения (иприт, диэтилнитрозамин, уретан и др.).
Некоторые лекарственные препараты также обладают мутагенным эффектом. Например, цитостатики, производные этиленимина, нитрозомочевина. Они повреждают ДНК в процессе репликации.
Химические мутагены могут вызывать нарушение мейоза, приводящее к нерасхождению хромосом, разрыву хромосом, точковым мутациям. Некоторые химические мутагены проходят через метаболическую систему организма самыми непредсказуемыми путями, превращаются в другие соединения. При этом они могут потерять свою мутагенную активность, или приобрести такие мутагенные свойства, которые отсутствовали у исходного соединения. Некоторые немутагенные химические вещества, включившись в обмен веществ, превращаются в мутагены. Например, цитостатик — циклофосфамид — не мутаген, но в организме млекопитающих превращается в высокомутагенное соединение.
Кроме мутагенов физической и химической природы, в окружающей среде имеются биологические факторы мутагенеза.
Вирусы оспы, кори, ветряной оспы, эпидемического паротита, гепатита, краснухи и др. способны вызывать разрывы хромосом. Вирусы могут усиливать темпы мутации клеток хозяина за счет подавления активности репарационных систем. Есть данные о возрастании числа хромосомных перестроек в клетках человека после пандемий, вызванных вирулентными вирусами.
Возникновение мутаций приводит к различным патологиям. Для предотвращения негативных последствий, связанных с действием различных мутагенных факторов среды, проводят мероприятия, снижающие вероятность возникновения мутаций. С этой целью используют вещества, называемые антимутагенными. В настоящее время выделено около 200 природных и синтетических соединений, обладающих антимутагенной активностью. Это аминокислоты (гистидин, метионин и др.), витамины (токоферол, каротин, ретинол, аскорбиновая кислота и др.), ферменты (оксидаза, каталаза и др.), интерферон и др.
Потребляемая пища содержит большое количество мутагенов и антимутагенов. Их соотношение зависит от способов обработки пищи, сроков ее хранения и т.д. Правильное питание — один из путей предотвращения вредного воздействия мутагенных факторов среды.
Как ухаживать за комнатной фуксией. почему у фуксии вянут и опадают листья вянут листья у фуксии что делать
2.1 Влияние ионизирующего облучения на живой организм
при действии физических мутагенов возникают так же, как и при действии
мутагенов химических. Вначале возникает первичное повреждение . Если оно не будет полностью исправлено в
результате ,
то при последующем репликативном синтезе будут возникать .
Специфика
(процесса возникновения )
при действии физических факторов связана с характером первичных повреждений , вызываемых ими.
Ионизирующее
излучение
– это поток заряженных или нейтральных частиц и квантов электромагнитного
излучения, прохождение которых через вещество приводит к ионизации и
возбуждению атомов или молекул среды.
Ионизирующее
излучение может вызвать мутации – внезапные естественные или вызванные
искусственно наследуемые изменения генетического материала, приводящие к
изменению тех или иных признаков организма.
Есть мутации спонтанные,
возникающие под влиянием природных факторов внешней среды или в результате
биохимических изменений в самом организме, и индуцированные, возникающие
под воздействием мутагенных факторов, например, ионизирующего излучения
химических веществ.
Мутации могут
быть прямыми, если их проявление приводит к отклонению от признаков так
называемого дикого типа и обратными, если они приводят к восстановлению
дикого типа.
Мутации в
половых клетках – генеративные – передаются следующим поколениям; мутации в
любых других клетках организма – соматические – наследуются только дочерними
клетками и оказывают воздействие лишь на тот организм, в котором возникли.
Ядерные
мутации затрагивают хромосомы ядра, цитоплазматические – генетический материал,
заключенный в цитоплазматических органоидах клетки – митохондриях, пластидах.
В зависимости
от характера изменений в генетическом материале различают точечные мутации,
геномные мутации и хромосомные аберрации (перестройки). Точечные мутации
представляют собой результат изменения последовательности нуклеотидов в
молекуле ДНК, являющейся носителем генетической информации и связаны с
добавлением, выпадением или перестановкой оснований в ДНК. Геномные мутации
связаны с изменением числа хромосом в клетке, кратным одинарному набору
хромосом, а также увеличением или уменьшением числа отдельных хромосом.
Радиоактивные
вещества могут воздействовать на организм человека внешне и внутренне. Внешнее
облучение характеризуется воздействием ионизирующего излучения извне и обусловлено
различной проникающей способностью частиц. Внутреннее облучение связано с
попаданием радиоактивного вещества внутрь человеческого организма с пищей, с
вдыхаемым воздухом или через открытую рану.
Воздействие
радиоактивного излучения на организм человека зависит от многих факторов и
определяется:
— скоростью
радиоактивного распада радионуклида;
— скоростью
выведения РВ из организма;
— типом
радиоактивного излучения;
Острые
последствия проявляются в первые несколько дней (недель) после облучения.
Отдаленные последствия – последствия, которые развиваются не сразу после
облучения, а спустя некоторое время.
Острая
лучевая болезнь возникает после тотального однократного внешнего равномерного
облучения. Между величиной поглощенной дозы в организме и средней продолжительностью
жизни существует строгая зависимость.
При воздействии ионизирующего излучения в дозах, не вызывающих
острую или хроническую лучевую болезнь, происходит изменениях в основных
регуляторных системах организма и функциональные изменения деятельности
основных физиологических систем чаще всего носят полисиндромный характер. Это
проявляется в развитии донозологических состояний, переходящих с ростом дозы к
клинической патологии.
В структуре неврологической заболеваемости особое место занимает
синдром вегетативной дистонии, повышения тревожности как устойчивой личностной
черты, отмечается ускорение перехода психофизиологических расстройств в стойкие
психосоматические.
При
дополнительном воздействии других неблагоприятных факторов существует вероятность
роста общесоматических заболеваний. Радиационный фактор выступает лишь как одно
из условий этого роста.
Отзывы из интернета
Примечания
- Banerjee S. K., Borden A., Christensen R. B., LeClerc J. E., Lawrence C. W. SOS-dependent replication past a single trans-syn T-T cyclobutane dimer gives a different mutation spectrum and increased error rate compared with replication past this lesion in uniduced cell // J. Bacteriol. — 1990. — 172. — P. 2105—2112.
- Jonczyk P., Fijalkowska I., Ciesla Z. Overproduction of the subunit of DNA polymerase III counteracts the SOS-mutagenic response of Esthetician coli // Proc. Nat. Acad. Sci. USA. — 1988. — 85. — Р. 2124—2127.
- Grebneva H. A. One of mechanisms of targeted substitution mutations formation at SOS-replication of double-stranded DNA containing cis-syn cyclobutane thymine dimers // Environ. Mol. Mutagen. — 2006. −47. — P. 733—745.
- Bresler S. E. Theory of misrepair mutagenesis // Mutat. Res. — 1975. — 29. — P. 467—472.
- ↑ Pham P., Bertram J. G, O’Donnell M., Woodgate R., Goodman M. F. A model for SOS-lesion-targeted mutations in Escherichia coli // Nature. — 2001. — 408. — P. 366—370.
- Taylor J.-S. New structural and mechanistic insight into the A-rule and the instructional and non-instructional behavior of DNA photoproducts and other lesions // Mutation. Res. — 2002. −510. — P. 55-70.
- Danilov V. I., Les A., Alderfer J. L. A theoretical study of the cis-syn pyrimidine dimers in the gas phase and water cluster and a tautomer — bypass mechanism for the origin of UV-induced mutations // J. Biomol. Struct. Dyn. — 2001. — 19. — P. 179—191.
- Gorb L., Podolyan Y., Dziekonski P., Sokalski W. A., Leszczynski J. Double-proton transfer in adenine-thymine and guanine-cytosine base pairs. A post-Hartree-Fock ab initio study // J. Am. Chem. Soc. — 2004. — 126. — P. 10119-10129.
- Полтев В. И., Шулюпина Н. В., Брусков В. И. Молекулярные механизмы правильности биосинтеза нуклеиновых кислот. Компьютерное изучение роли полимераз в образовании неправильных пар модифицированными основаниями // Молек. биол. — 1996. — 30. — С. 1284—1298.
- Cannistraro V. J., Taylor J. S. Acceleration of 5-methylcytosine deamination in cyclobutane dimers by G and its implications for UV-induced C-to-T mutation hotspots // J. Mol. Biol. — 2009. — 392. — P. 1145—1157.
- Тарасов В. А. Молекулярные механизмы репарации и мутагенеза. — М.: Наука, 1982. — 226 с.
- Friedberg E. C., Walker G. C., Siede W. DNA repair and mutagenesis. — Washington: ASM Press, DC, 1995.
- Ауэрбах Ш. Проблемы мутагенеза. — М.: Мир, 1978. — 463 с.
- Friedberg E. C., Walker G. C., Siede W., Wood R. D., Schultz R. A., Ellenberger T. DNA repair and mutagenesis. — part 3. Washington: ASM Press. — 2006. 2nd ed.
- Levine J. G., Schaaper R. M., De Marini D. M. Complex frameshift mutations mediated by plasmid pkm 101: Mutational mechanisms deduced mutation spectra in Salmonella // Genetics. — 1994. — 136. — P. 731—746.
- Wang C.-I., Taylor J.-S. In vitro evidence that UV-induced frameshift and substitution mutations at T tracts are the result of misalignment-mediated replication past a specific thymine dimer // Biochemistry — 1992. — 31. — P. 3671-3681.
- Maor-Shoshani A., Reuven N. B., Tomer G., Livneh Z. Highly mutagenic replication by DNA polymerase V (UmuC) provides a mechanistic basis for SOS untargeted mutagenesis // Proc. Natl. Acad. Sci. USA — 2000. — 97. — P. 565—570.
- Little J. B., Gorgojo L., Vetrovs H. Delayed appearance of lethal and specific gene mutations in irradiated mammalian cells // Int. J. Radiat. Oncol. Biol. Phys. — 1990. — 19. — P. 1425—1429.
- Niwa O. Radiation induced dynamic mutations and transgenerational effects // J. Radiation Research. — 2006. — 47. — P. B25-B30.
- Самигуллина Н. С. Практикум по селекции и сортоведению плодовых и ягодных культур: Учебное издание. — Мичуринск: Мичуринский государственный аграрный университет, 2006. — 197 с.
Выявление источников мутагенов в окружающей среде (косвенно) и оценка возможных последствий их влияния на собственный организм
Источники мутагенов способны оказывать косвенное влияние на окружающую среду и здоровье человека. Рассмотрим основные категории данных опасных соединений и особенности их воздействия:
| В быту | Выраженной мутагенностью обладают красители для волос, бытовая химия и отдельные продукты питания. Для защиты организма от этих веществ необходимо меньше употреблять в пищу «сомнительных» продуктов питания, а моющие средства использовать в перчатках. |
| На производстве | Мутагенное воздействие на производстве способно проникать в человеческий организм посредством пищеварительного тракта, кожи и легких. Наиболее опасными признаны следующие соединения: стирол, эпоксидные смолы, эпихлоргидрин, хлоропрен и винилхлорид. Поэтому люди, работающие с такими веществами, одевают специальные средства защиты и раньше уходят на заслуженный отдых. |
Смотри также:
- Виды мутаций и их причины. Значение изменчивости в жизни организмов и в эволюции
- Значение генетики для медицины. Наследственные болезни человека, их причины, профилактика
- Селекция, ее задачи и практическое значение
Как мариновать репу
У человека была измерена частота мутаций
В течение многих лет мы могли оценивать уровень мутаций человека только при помощи косвенных методов. Первая попытка их подсчёта предпринята в 1930 г. Дж. Б. С. Холдейном в работе по гемофилии. Теперь с помощью быстрого и дешёвого способа секвенирования целых геномов мы можем непосредственно измерять частоту мутаций человека, сравнивая геномы родителей и потомства.
Недавнее исследование семьи из Исландии говорит о том, что у каждого человека в течение жизни происходит примерно 70 мутаций и 76% из них абсолютно новые. Они наблюдаются в отцовском геноме, и их количество увеличивается с возрастом отца. Другие методы, включая оригинальный метод Холдейна, нашли причину отцовского уклона. Вероятнее всего это отражение популяции стволовых клеток, которые дают начало сперматозоидам, мутирующим с возрастом.
Вставки и удаления не менее 50 оснований встречаются с частотой примерно от 1/5 до 1/10 новых мутаций. Удаления и дублирования происходят гораздо медленнее (1 новая мутация на 42 новорождённых). Некоторые из них – мобильные генетические элементы, засоряющие наш геном, могут активно передвигаться. Скорость вставки мобильного элемента равна 1 на 20 родов.
Если бы со временем в генах не происходило никаких изменений, то эволюции бы не случилось. Однако слишком много изменений вредны для особи. Нужно соблюдение хрупкого баланса между количеством новых вариаций, возникающих у всего вида, и здоровьем отдельных особей.
Типы мутагенных агентов
Агенты, которые вызывают мутации в генетическом материале, очень разнообразны по своей природе. Сначала мы рассмотрим классификацию мутагенов и приведем примеры каждого типа, а затем объясним различные способы, которыми мутагены могут вызывать изменения в молекуле ДНК..
Химические мутагены
Мутагены химической природы включают следующие классы химических веществ: акридины, нитрозамины, эпоксиды и другие. Существует подклассификация этих агентов в:
Аналогичные основы
Молекулы, которые имеют структурное сходство с азотистыми основаниями, обладают способностью вызывать мутации; l 5-бромурацил и 2-аминопурин являются наиболее распространенными.
Агенты, которые реагируют с генетическим материалом
Азотистая кислота, гидроксиламин и ряд алкилирующих агентов реагируют непосредственно в основаниях, которые составляют ДНК, и могут переходить из пурина в пиримидин и наоборот.
Интерстициальные агенты
Существует целый ряд молекул, таких как акридины, бромид этидия (широко используемые в лабораториях молекулярной биологии) и профлавин, которые имеют плоскую молекулярную структуру и способны проникать в цепь ДНК..
Окислительные реакции
Нормальный метаболизм клетки имеет в качестве вторичного продукта ряд активных форм кислорода, которые повреждают клеточные структуры, а также генетический материал..
Физические мутагены
Второй тип мутагенных агентов — физические. В этой категории мы находим различные типы излучения, которые влияют на ДНК.
Наконец, у нас есть биологические мутанты. Это организмы, которые могут вызывать мутации (в том числе аномалии на уровне хромосом) в вирусах и других микроорганизмах.
Искусственный мутагенез
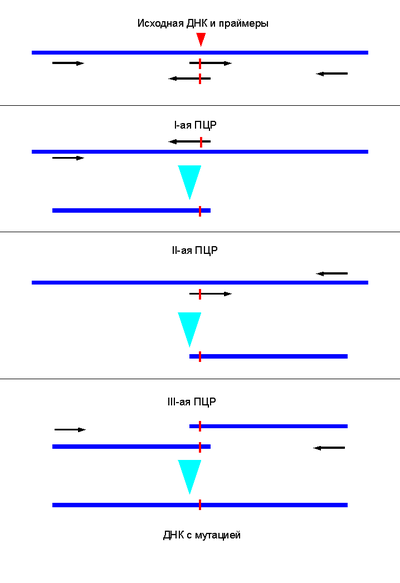 Сайт-направленный мутагенез. Синтезируют пару праймеров, несущих мутацию, и пару праймеров, комплементарных концам нужного фрагмента ДНК. В ходе первых двух реакций образуются фрагменты ДНК с мутацией, которые объединяют в третьей реакции. Полученный фрагмент вставляют в нужную генно-инженерную конструкцию.
Сайт-направленный мутагенез. Синтезируют пару праймеров, несущих мутацию, и пару праймеров, комплементарных концам нужного фрагмента ДНК. В ходе первых двух реакций образуются фрагменты ДНК с мутацией, которые объединяют в третьей реакции. Полученный фрагмент вставляют в нужную генно-инженерную конструкцию.
Искусственный мутагенез широко используют для изучения белков и улучшения их свойств (направленной эволюции (англ.)).
Ненаправленный мутагенез
Методом ненаправленного мутагенеза в последовательность ДНК вносятся изменения с определенной вероятностью. Мутагенными факторами (мутагенами) могут быть различные химические и физические воздействия — мутагенные вещества, ультрафиолет, радиация. После получения мутантных организмов производят выявление (скрининг) и отбор тех, которые удовлетворяют цели мутагенеза. Ненаправленный мутагенез более трудоемок и его проведение оправдано, если разработана эффективная система скрининга мутантов.
Направленный мутагенез
В направленном (сайт-специфическом) мутагенезе изменения в ДНК вносятся в заранее известный сайт (DNA binding site). Для этого синтезируют короткие одноцепочечные молекулы ДНК (праймеры), комплементарные целевой ДНК за исключением места мутации.
Мутагенез по Кункелю
Для бактериальной плазмиды (внехромосомной кольцевой ДНК) получают уридиновую матрицу, то есть такую же молекулу, в которой остатки тимина заменены на урацил. Праймер отжигают на матрице, проводят его достройку in vitro с помощью полимеразы до кольцевой ДНК, комплементарной уридиновой матрице. Двухцепочечной гибридной ДНК трансформируют бактериальные клетки, внутри клетки уридиновая матрица разрушается как чужеродная, и на мутантной одноцепочеченой кольцевой ДНК достраивается вторая цепь. Эффективность такого способа мутагенеза менее 100 %.
Мутагенез с помощью ПЦР
Полимеразная цепная реакция позволяет проводить сайт-направленный мутагенез с использованием пары праймеров, несущих мутацию, а также случайный мутагенез. В последнем случае ошибки в последовательность ДНК вносятся полимеразой в условиях, понижающих её специфичность.
Проблема случайности мутаций
В 40-е годы среди микробиологов была популярна точка зрения, согласно которой мутации вызываются воздействием фактора среды (например, антибиотика), к которому они позволяют адаптироваться. Для проверки этой гипотезы был разработан флуктуационный тест и метод реплик.
Флуктуационный тест Лурии-Дельбрюка заключается в том, что небольшие порции исходной культуры бактерий рассеивают в пробирки с жидкой средой, а после нескольких циклов делений добавляют в пробирки антибиотик. Затем (без последующих делений) на чашке Петри с твердой средой высевают выживших устойчивых к антибиотику бактерий. Тест показал, что число устойчивых колоний из разных пробирок очень изменчиво — в большинстве случаев оно небольшое (или нулевое), а в некоторых случаях очень высокое. Это означает, что мутации, вызвавшие устойчивость к антибиотику, возникали в случайные моменты времени как до, так и после его воздействия.
Метод реплик заключается в том, что с исходной чашки Петри, где на твердой среде растут колонии бактерий, делается отпечаток на ворсистую ткань, а затем с ткани бактерии переносятся на несколько других чашек, где рисунок их расположения оказывается тем же, что на исходной чашке. После воздействия антибиотиком на всех чашках выживают колонии, расположенные в одних и тех же точках. Высевая такие колонии на новые чашки, можно показать, что все бактерии внутри колонии обладают устойчивостью.
Таким образом, обоими методами было доказано, что «адаптивные» мутации возникают независимо от воздействия того фактора, к которому они позволяют приспособиться, и в этом смысле мутации случайны. Однако несомненно, что возможность тех или иных мутаций зависит от генотипа и канализована предшествующим ходом эволюции (см. Закон гомологических рядов в наследственной изменчивости).
Кроме того, закономерно различается частота мутирования разных генов и разных участков внутри одного гена. Также известно, что высшие организмы используют «целенаправленные» (то есть происходящие в определенных участках ДНК) мутации в механизмах иммунитета[источник не указан 1531 день]. С их помощью создаётся разнообразие клонов лимфоцитов, среди которых в результате всегда находятся клетки, способные дать иммунный ответ на новую, неизвестную для организма болезнь. Подходящие лимфоциты подвергаются положительной селекции, в результате возникает иммунологическая память. (В работах Юрия Чайковского говорится и о других видах направленных мутаций.)
Мутагенные факторы. Примеры
К мутагенам физического происхождения можно отнести источники воздействие ультрафиолета, аномально высокие или низкие температуры, влажность.

К примеру, волны ультрафиолетового излучения, имеющие длину свыше 260 нм, поглощаются клеткой листа растения и вызывают образование в ней нехарактерных пиримидиновых димеров (соединений в цепи ДНК), которые, в свою очередь, становятся причиной ошибок в считывании генетического материала. В результате новые клетки получают с «неправильной» структурой.
Многие химические вещества относятся к мутагенам и промутагенам. Примерами могут служить активные формы кислорода, нитраты и нитриты, некоторые металлы, лекарства и те вещества, которых до появления человечества в природе не существовало (бытовая химия, пищевые добавки и консерванты).
К примеру, беременная женщина может не знать о своем положении и принимать некоторые антибиотики, опасные для плода. В результате у ребенка могут развиться вызванные мутациями.
Результатом воздействия таких биологических агентов на клетку является процесс, который называется инфекционным мутагенезом. К примеру, бактерия Helicobacter pylori, живущая в кишечнике и желудке человека, может вызывать воспалительные процессы на слизистой. Воспаление изменяет нормальный ход окислительно-восстановительных процессов в поврежденных клетках, что меняет и структуру генетического материала в них. Нарушаются процессы восстановления ДНК и ход нормального деления молекулы. Результат — мутации.

Адаптивная ценность
Большая часть новых мутантов отличается существенно низкой жизнеспособностью, чем дикий/нормальный тип. При этом она выражается в различной степени: от субвитального едва заметного до полулетального и летального состояния. При анализе жизнеспособности мутантов мухи дрозофилы, появившихся при изменениях в Х-хромосоме, у 90% особей она была ниже, чем у нормальных. У 10% отмечалось супервитальное состояние – повышенная жизнеспособность. В целом же адаптивная ценность появившихся мутантов, как правило, понижена. Она характеризуется функциональной полезностью морфологических признаков и плодовитостью, физиологической жизнеспособностью.
Роль мутаций в эволюции
При существенном изменении условий существования те мутации, которые раньше были вредными, могут оказаться полезными. Таким образом, мутации являются материалом для естественного отбора. Так, мутанты-меланисты (темноокрашенные особи) в популяциях березовой пяденицы в Англии впервые были обнаружены учеными среди типичных светлых особей в середине XIX века. Темная окраска возникает в результате мутации одного гена. Бабочки проводят день на стволах и ветвях деревьев, обычно покрытых лишайниками, на фоне которых светлая окраска является маскирующей. В результате промышленной революции, сопровождающейся загрязнением атмосферы, лишайники погибли, а светлые стволы берез покрылись копотью. В результате к середине XX века (за 50-100 поколений) в промышленных районах темная морфа почти полностью вытеснила светлую. Было показано, что главная причина преимущественного выживания чёрной формы — хищничество птиц, которые избирательно выедали светлых бабочек в загрязненных районах.
Если мутация затрагивает «молчащие» участки ДНК либо приводит к замене одного элемента генетического кода на синонимичный, то она обычно никак не проявляется в фенотипе (проявление такой синонимичной замены может быть связано с разной частотой употребления кодонов). Однако методами генного анализа такие мутации можно обнаружить. Поскольку чаще всего мутации происходят в результате естественных причин, то в предположении, что основные свойства внешней среды не менялись, получается, что частота мутаций должна быть примерно постоянной. Этот факт можно использовать для исследования филогении — изучения происхождения и родственных связей различных таксонов, в том числе и человека. Таким образом, мутации в молчащих генах служат для исследователей «молекулярными часами». Теория «молекулярных часов» исходит также из того, что большинство мутаций нейтрально, и скорость их накопления в данном гене не зависит или слабо зависит от действия естественного отбора и потому остается постоянной в течение длительного времени. Для разных генов эта скорость, тем не менее, будет различаться.
Исследование мутаций в митохондриальной ДНК (наследуется по материнской линии) и в Y-хромосомах (наследуется по отцовской линии) широко используется в эволюционной биологии для изучения происхождения рас и антропологических типов, реконструкции биологического развития человечества.
Мутация генов
Генные мутации – это наиболее распространённый тип наследственной изменчивости, они связаны с деформацией первичной структуры ДНК в пределах одного гена, происходят на молекулярном уровне организации живого. Возникают они в процессе удвоения ДНК. Происходит это нечасто, примерно одна мутация на 10-100 тыс. копий. Но в генотипах они постепенно накапливаются, создавая разнообразие всего живого. Именно благодаря таким мутациям возникли разные аллели одного гена.
Мутация, при которой изменяется одна пара оснований, называется точечной или однонуклеотидной. Нуклеотид может быть заменён на другой, добавлен или удалён.
Мутация: замена азотистых оснований
Замена одной пары оснований в ДНК на другую называется мутацией замещения. Из-за вырожденной природы генетического кода замена основания может не привести к замещению закодированной аминокислоты. Если окажется, что новый триплет кодирует ту же аминокислоту, то мутация будет молчащей.
Часто при замене основания меняется кодируемая аминокислота в белке, получаемом при транскирипции мутантного гена. Замены делятся на две группы:
- транзакции;
- трансверсии.
Транзакция не заменяет типа оснований в гене, пиримидин замещается пиримидином, а пурин – пурином. Напротив, трансверсия изменяет тип оснований в паре (приримидин – пурин). Различные заболевания вызваны трансверсией, в том числе серповидно-клеточная анемия.

Бессмысленные (нонсенс-мутации)
Особая категория замены оснований вызывает смену транскрибируемого (кодирующего белок) кодона в стоп-кодон. Он приводит к преждевременному прекращению трансляции и к усечению белка. Насколько коротким получится белок зависит от того, где в гене установится стоп-кодон.
Может быть и наоборот, когда стоп-кодон в результате мутации становится активным геном и способствует синтезированию необычно длинных молекул белка.
Мутация сдвига рамки считывания
Добавление или удаление пары комплементарных оснований имеет гораздо большие последствия, чем замена одного основания на другое. Такие нарушения сдвигают рамки считывания в мРНК ниже от мутации. Это связано с тем, что считывание информации происходит триплетами без знаков препинания внутри тройки. Обычно такие мутации приводят к полному выключению функционирования гена. Этот тип деформаций был использован Криком и Бреннером для того, чтобы сделать вывод о триплетной природе генетического кода.
Изменение рамки считывания в гене на ранней стадии означает изменение части белка. Смена кадров также может привести к преждевременному завершению транскрипции, так как 3 из 64 кодонов являются стоп-кодонами.
Генная мутация: удвоение триплета
Учитывая долгую историю молекулярной генетики и относительно короткое время молекулярного анализа у человека, удивительно, что новый вид мутации был обнаружен у людей.
Один из первых генов, связанных с нарушением у человека – болезни Хантингтона – был мутантным. При этом ген, кодирующий белок хантингтин, содержит повторяющийся триплет. Болезнь Хантингтона — дегенеративное расстройство нервной системы, которое приводит к совершению неконтролируемых движений, утрате интеллектуальных способностей и эмоциональному возбуждению. При болезни Хантингтона белок, кодируемый мутировавшим геном, убивает нервные клетки в определённых частях мозга, следствием чего становится гибель нервов, которые регулируют движения.
Болезнь Хантингтона обнаруживается во взрослом возрасте (в то время как остальные генетически обусловленные болезни обычно проявляются в детстве), и поэтому те, кто болен ею, часто успевают произвести на свет детей. Мутация гена, обуславливающая болезнь Хантингтона, передается как через мужские, так и через женские хромосомы, и поэтому существует очень большая вероятность, что ребёнок обязательно унаследует заболевание от больного родителя.
Теперь известно не менее 20 заболеваний, связанных с такой мутацией. Такой тип аномалий пока был найден только у людей и мышей. Предполагают, что он характерен только для позвоночных или даже исключительно для млекопитающих. Например, у дрозофилы её не обнаруживали никогда.
Расширение триплета может происходить в области кодирования или транскрибирования. В случае болезни Хантингтона единица повтора находится в кодирующей области триплета, кодирующего глутамин, и расширение приводит к полиглутаминовой области в белке. В случае синдрома Мартина-Белла наследственная форма умственной отсталости вызвана повтором в некодирующей части ДНК.













